Respon Imun - Ketika
mikroba masuk ke dalam tubuh manusia, mikroba tersebut akan melewati 3 lapis
pertahanan sistem imun.
Pertahanan lapis pertama berisi sistem imun non-spesifik terutama
fisik/mekanis, biokimia, dan humoral. Pertahanan ini akan mencegah masuknya
mikroba masuk ke dalam tubuh. Pertahanan lapis kedua berisi sistem imun
non-spesifik khususnya yang selular. Pertahanan selular ini nantinya akan
mencegah mikroba yang berhasil masuk ke dalam tubuh dengan menghancurkannya.
Pertahanan ketiga adalah sistem imun spesifik yang telah dibahas di atas. Ini
akan menangani mikroba yang masih belum ditangani oleh sistem imun non-spesifik.
Faktor yang Mempengaruhi Sistem Imun
Beberapa
faktor yang mempengaruhi sistem imun,
yaitu :
1. Usia
2. Jenis
kelamin
3. Lingkungan
Mekanisme Pertahanan Tubuh
Sistem
kekebalan tubuh yang sehat merupakan kekebalan yang dapat membedakan antara
bagian tubuh dari sistem itu sendiri dan benda asing yang masuk ke dalam tubuh.
Secara garis besar, sistem imun menurut sel tubuh dibagi menjadi sistem imun
humoral dan sistem imun seluler. Sistem imun humoral terdiri atas antibodi dan
cairan yang disekresikan organ tubuh tubuh (saliva, air mata, serum, keringat,
asam lambung, pepsin, dan lain-lain). Sedangkan sistem imun dalam bentuk
seluler berupa makrofag, limfosit, dan neutrofil yang berada di dalam sel.
Tubuh
manusia mempunyai banyak sekali mekanisme pertahanan yang terdiri dari berbagai
macam sistem imun yaitu organ limfoid (thymus, lien, sumsum tulang) beserta
sistem limfatiknya. Jantung, hati, ginjal, dan paru-paru juga termasuk dalam
mekanisme pertahanan tubuh. Sistem limfatik baru akan dikatakan mengalami
gangguan jika muncul tonjolan yang membesar dibandingkan keadaan biasanya. Hal
ini dikarenakan kelenjar limfe sedang berpasangan melawan kuman yang masuk
dalam tubuh. Organ limfoid seperti thymus sendiri mempunyai
tanggungjawab dalam pembentukan sel T. Kelenjar thymus sangat penting
bagi bayi yang baru lahir, karena bayi yang tidak memiliki
kelenjarthymus akan mempunyai sistem imun yang buruk.
Leukosit (sel darah putih) dihasilkan
oleh thymus, lien dan sumsum tulang belakang.
Leukosit
bersirkulasi di dalam tubuh melalui pembuluh limfe dan pembuluh darah, sehingga
sistem imun bekerja terkoordinasi baik memonitor tubuh dari kuman maupun
substansi lain yang bisa menyebabkan permasalahan dalam tubuh. Leukosit pada
umumnya memiliki dua tipe, yaitu fagosit yang bertugas memakan organisme yang
masuk ke dalam tubuh dan limfosit yang bertugas mengingat dan mengenali yang
masuk ke dalam tubuh serta membantu tubuh menghancurkan benda asing tersebut.
Sel lainnya adalah netrofil, yang bertugas melawan bakteri. Kadar netrofil bisa
dijadikan indikator adanya infeksi dari bakteri.
Limfosit
terdiri dari dua tipe, yaitu limfosit B dan Limfosit T. Limfosit dihasilkan
oleh sumsum tulang belakang. Limfosit yang berada di dalam sumsum tulang
belakang jika matang menjadi limfosit sel B, atau jika meninggalkan sumsum
tulang belakang menuju kelenjar thymus menjadi limfosit T.
Limfosit B
dan T mempunyai fungsi yang berbeda dimana limfosit B berfungsi untuk mencari
target dan mengirimkan “tentara” untuk mengunci keberadaan benda asing. Benda
asing yang telah diidentifikasi oleh sel B kemudian akan dihancurkan oleh sel
T. Jika terdapat antigen (benda asing yang masuk ke dalam tubuh) terdeteksi,
maka beberapa tipe sel bekerjasama untuk mencari tahu sel yang akan memberikan
respon. Sel-sel ini memicu limfosit B untuk memproduksi antibodi, suatu protein
khusus yang mengarahkan kepada suatu antigen spesifik. Antibodi sendiri bisa
menetralisir toksin yang diproduksi dari berbagai macam organisme, dan juga
antibodi bisa mengaktivasi kelompok protein protein yang disebut komplemen yang
merupakan bagian dari sistem imun dan membantu menghancurkan bakteri, virus,
mikroorganisme patogen, ataupun sel yang terinfeksi.
Sistem Kekebalan Tubuh Pada Manusia
Mekanisme Imunitas terhadap Antigen yang Berbahaya
Beberapa mekanisme pertahanan tubuh
dalam mengatasi agen yang berbahaya di lingkungannya yaitu:
·
Pertahanan fisik dan kimiawi: kulit, sekresi
asam lemak dan asam laktat melalui kelenjar keringat dan sebasea (kelenjar
berbentuk kantong kecil yang terletak di dermis), sekresi lendir, pergerakan
silia, sekresi air mata, air liur, urin, asam lambung serta lisozim dalam air
mata. Simbiosis dengan bakteri flora normal yang memproduksi zat yang dapat
mencegah invasi mikroorganisme seperti laktobasilus pada epitel organ.
·
Innate immunity
·
Imunitas
spesifik yang didapat.
Respon Imune Innate
Respon ini
merupakan mekanisme pertahanan tubuh non-spesifik yang mencegah masuk dan
menyebarnya mikroorganisme dalam tubuh serta mencegah terjadinya kerusakan
jaringan. Ada beberapa komponen innate immunity, yaitu :
·
Pemusnahan bakteri intraselular oleh sel
poli-morfonuklear (PMN) dan makrofag.
·
Aktivasi komplemen melalui jalur alternatif.
·
Degranulasi sel mast yang melepaskan mediator
inflamasi.
·
Protein fase akut: C-reactive
protein (CRP) yang mengikat mikroorganisme, selanjutnya terjadi aktivasi komplemen
melalui jalur klasik yang menyebabkan lisis mikroorganisme.
·
Produksi interferon alfa (IFN-α) oleh leukosit
dan interferon beta (IFN-β) oleh fibroblast yang mempunyai efek antivirus.
·
Pemusnahan mikroorganisme ekstraselular oleh
sel natural killer (sel NK) melalui pelepasan granula yang mengandung
perforin.
·
Pelepasan mediator eosinofil
seperti major basic protein (MBP) dan protein kationik yang
dapat merusak membran parasit.
Respon Imunitas Spesifik
Bila
mikroorganisme dapat melewati pertahanan nonspesifik/innate immunity, maka
tubuh akan membentuk mekanisme pertahanan yang lebih kompleks dan spesifik.
Mekanisme imunitas ini memerlukan pengenalan terhadap antigen lebih dulu.
Mekanisme imunitas spesifik ini terdiri dari imunitas humoral, yaitu produksi
antibodi spesifik oleh sel limfosit B (Tdependent dan non
T dependent) dan mekanisme Cell mediated immunity (CMI). Sel
limfosit T berperan pada mekanisme imunitas ini melalui produksi sitokin serta
jaringan interaksinya dan sel sitotoksik matang di bawah pengaruh interleukin 2
(IL-2) dan interleukin 6 (IL-6).
Presentasi Antigen
Respons
imun tubuh dipicu oleh masuknya antigen/mikroorganisme ke dalam tubuh dan
dihadapi oleh sel makrofag yang selanjutnya akan berperan sebagai antigen
presenting cell (APC). Sel itu akan menangkap sejumlah kecil antigen dan
diekspresikan ke permukaan sel yang dapat dikenali oleh sel limfosit Th atau
T helper. Sel Th ini akan teraktivasi dan (selanjutnya sel Th ini)
akan mengaktivasi limfosit lain seperti sel limfosit B atau sel limfosit T
sitotoksik. Sel T sitotoksik ini kemudian berpoliferasi dan mempunyai fungsi
efektor untuk mengeliminasi antigen. Sel limfosit dan sel APC bekerja sama
melalui kontak langsung atau melalui sekresi sitokin regulator. Sel-sel ini
dapat juga berinteraksi secara simultan dengan sel tipe lain atau dengan
komponen komplemen, kinin atau sistem fibrinolitik yang menghasilkan aktivasi
fagosit, pembekuan darah atau penyembuhan luka. Respon imun dapat bersifat
lokal atau sistemik dan akan berhenti bila antigen sudah berhasil dieliminasi
melalui mekanisme kontrol.
Peran Major Histocompatibility Complex (MHC)
Respon
imun sebagian besar antigen hanya dimulai bila antigen telah ditangkap dan
diproses serta dipresentasikan oleh sel APC. Oleh karena itu, sel T hanya
mengenal imunogen yang terikat pada protein MHC pada permukaan sel lain.
terdapat 2 kelas MHC yaitu:
Protein
MHC kelas I. Diekspresikan oleh semua tipe sel somatik dan digunakan untuk
presentasi antigen kepada sel TCD8 yang sebagian besar adalah sel sitotoksik.
Hampir sebagian besar sel mempresentasikan antigen ke sel T sitotoksik (sel Tc)
serta merupakan target/sasaran dari sel Tc tersebut. MHC kelas I digunakan
ketika merepson infeksi virus.
Protein
MHC kelas II. Diekspresikan hanya oleh makrofag dan beberapa sel lain untuk
presentasi antigen kepada sel TCD4 yang sebagian besar adalah sel
T helper (Th). Aktivasi sel Th ini diperlukan untuk respon imun yang
sesungguhnya dan sel APC dengan MHC kelas II merupakan poros penting dalam
mengontrol respon imun tersebut. MHC kelas II digunakan ketika merespon infeksi
bakteri.
T Helper 1 (Th1) dan T Helper 2 (Th2)
Sel-sel T
berperan sebagai penghantar imunitas yang dimediasi sel dalam respon imun
adaptif yang digunakan untuk mengontrol patogen intraseluler serta meregulasi
respon sel B, termasuk aktivasi sel imun lainnya dengan pelepasan sitokin (Uzel
2000). Terdapat dua subset utama limfosit yang dibedakan dengan keberadaan
molekul (petanda) permukaan CD4 dan CD8. Limfosit T yang mengekspresikan CD4
juga dikenal sebagai sel T helper, penghasil sitokin terbanyak. Subset ini
dibagi lagi menjadi Th1 dan Th2, dan sitokin yang dihasilkan disebut sebagai
sitokin tipe Th1 dan sitokin tipe Th2. Sitokin tipe Th1 cenderung menghasilkan
respon proinflamatori yang bertanggung jawab terhadap killingparasit
intraseluler dan mengabadikan respon autoimun. Sitokin tipe Th1 terdiri dari
interferon gamma, interleukin-2, serta limfotoksin-α yang merangsang imunitas
tipe 1, ditandai aktivitas fagositik yang kuat.
Respon
proinflamatori yang berlebihan akan mengakibatkan kerusakan jaringan yang tidak
terkontrol. Tubuh mempunyai suatu mekanisme untuk menetralkan aksi
mikrobisidal berlebih yang dimediasi Th1 ini, yaitu dengan respon Th2.
Sitokin yang termasuk dalam mekanisme Th2 ini adalah interleukin 4, 5, 9, dan
13, yang disertai IgE dan respon eosinofilik dalam atopi, dan juga
interleukin-10, dengan respon yang lebih bersifat anti-inflamatori. Imunitas
tipe 2 yang distimulasi Th2 ditandai dengan kadar antibodi tinggi (Berger 2000).
Bagi kebanyakan infeksi, imunitas tipe 1 bersifat protektif, sedang respon tipe
2 membantu resolusi inflamasi yang dimediasi sel. Stres sistemik yang berat,
imunosupresi, atau inokulasi mikrobial yang berlebihan (overwhelming)
mengakibatkan sistem imun meningkatkan respon tipe 2 terhadap infeksi
yang seharusnya dikendalikan oleh imunitas tipe 1 (Spellberg 2001).
Kemungkinan prekursor sel-T penolong akan menjadi sel tipe 1 atau tipe 2
tergantung pada beberapa faktor, yaitu dilihat dari sudut pandang patogen
seperti sifat dan kuantitas patogen, route infeksi, pengaruh komponen
imunomodulator dan infeksi bersamaan, serta faktor pejamu termasuk predisposisi
genetik, jumlah sel-T yang merespon, kompleks histokompatiliti mayor haplotype
individu, sifat sel yang mempresentasikan antigen, serta lingkungan sitokin
sel-T selama dan pasca aktivasi (Nahid 1999).
Mekanisme Respon Tubuh terhadap Serangan Mikroba
Respons
tubuh terhadap serangan mikroba dapat terjadi dalam beberapa jenjang tahapan.
Tahapan pertama bersifat nonspesifik atau innate, yaitu berupa
respons inflamasi. Tahapan kedua bersifat spesifik dan adaptif, yang diinduksi
oleh komponen antigenik mikroba. Tahapan terakhir adalah respon peningkatan dan
koordinasi sinergistik antara sel spesifik dan nonspesifik yang diatur oleh
berbagai produk komponen respon inflamasi, seperti mediator kimia. Sistem
kekebalan adalah sistem pertahanan manusia sebagai perlindungan terhadap
infeksi dari makromolekul asing atau serangan organisme, termasuk virus,
bakteri, protozoa dan parasit. Sistem kekebalan dalam tubuh juga berperan dalam
perlawanan terhadap protein tubuh dan molekul lain seperti yang terjadi pada
autoimunitas, dan melawan sel yang teraberasi menjadi tumor.
Kemampuan
sistem kekebalan untuk membedakan komponen sel tubuh dari komponen patogen
asing akan menopang amanat yang diembannya guna merespon infeksi patogen, baik
yang berkembang biak di dalam sel tubuh (intraseluler) seperti misalnya virus,
maupun yang berkembang biak di luar sel tubuh (ekstraseluler) sebelum
berkembang menjadi penyakit. Meskipun demikian, sistem kekebalan mempunyai sisi
yang kurang menguntungkan. Pada proses peradangan, penderita dapat merasa tidak
nyaman oleh karena efek samping yang dapat ditimbulkan sifat toksik senyawa
organik yang dikeluarkan sepanjang proses perlawanan berlangsung. Pertahanan
awal terhadap organisme asing adalah jaringan terluar dari tubuh yaitu kulit,
yang memiliki banyak sel termasuk makrofag dan neutrofil yang siap memfagosit
organisme lain pada saat terjadi penetrasi pada permukaan kulit, dengan tidak
dilengkapi oleh antibodi. Pertahanan yang kedua adalah kekebalan tiruan.
Walaupun
sistem pada kedua pertahanan mempunyai fungsi yang sama, terdapat beberapa
perbedaan yang nyata, antara lain :
·
sistem kekebalan tiruan tidak dapat terpicu
secepat sistem kekebalan turunan
·
sistem kekebalan tiruan hanya merespon
imunogen tertentu, sedangkan sistem yang lain merespon nyaris seluruh antigen.
·
sistem kekebalan tiruan menunjukkan kemampuan
untuk “mengingat” imunogen penyebab infeksi dan reaksi yang lebih cepat saat
terpapar lagi dengan infeksi yang sama. Sistem kekebalan turunan tidak
menunjukkan kemampuan immunological memory.
Imunologi Toleransi Terhadap Antigen bakteri
Toleransi
adalah properti dari host dimana ada pengurangan imunologis spesifik dalam
respon imun terhadap antigen tertentu. Toleransi ke Antigen bakteri tidak
melibatkan kegagalan umum dalam respon imun tetapi kekurangan tertentu dalam
kaitannya dengan antigen dari bakteri tertentu. Jika ada respon kekebalan
yang tertekan terhadap antigen yang relevan dari parasit, proses infeksi
difasilitasi. Toleransi dapat melibatkan baik AMI (Antibody-Mediated Immunity)
atau CMI (Cell Mediated Immunity) atau kedua lengan dari respon imunologi.
Toleransi terhadap suatu Antigen dapat timbul dalam berbagai cara, tetapi tiga
yang mungkin relevan dengan infeksi bakteri.
1. Paparan
Antigen Janin terpapar Antigen. Jika janin terinfeksi pada tahap tertentu
dari perkembangan imunologi, mikroba Antigen dapat dilihat sebagai “diri”,
dengan demikian menyebabkan toleransi (kegagalan untuk menjalani respon
imunologi) ke Antigen yang dapat bertahan bahkan setelah kelahiran.
2. High
persistent doses of circulating Antigen. Toleransi terhadap bakteri atau salah
satu produknya mungkin timbul ketika sejumlah besar antigen bakteri yang
beredar dalam darah menyebabkan sistem kekebalan menjadi kewalahan.
3. Molecular
mimicry. Jika Antigen bakteri sangat mirip dengan “antigen” host normal, respon
kebal terhadap Antigen ini mungkin lemah memberikan tingkat toleransi.
Kemiripan antara Antigen bakteri dan host Antigen disebut sebagai mimikri
molekuler. Dalam hal ini determinan antigenik dari bakteri sangat erat
terkait kimiawi untuk host komponen jaringan yang sel-sel imunologi tidak dapat
membedakan antara dua dan respon imunologi tidak dapat ditingkatkan.
Beberapa kapsul bakteri tersusun dari polisakarida (hyaluronic acid, asam
sialic) sehingga mirip dengan host polisakarida jaringan yang tidak imunogenik.
Antibodi yang diserap oleh Antigen bakteri Larut
Beberapa
bakteri dapat membebaskan komponen antigen permukaan dalam bentuk yang larut ke
dalam cairan jaringan. Antigen ini larut dan dapat menggabungkan dengan
“menetralisir” antibodi sebelum mereka mencapai sel-sel bakteri.
Misalnya, sejumlah kecil endotoksin (LPS) dapat dilepaskan ke cairan sekitarnya
oleh bakteri Gram-negatif. Otolisis bakteri Gram-negatif atau Gram-positif
dapat melepaskan komponen antigen permukaan dalam bentuk yang
larut.Streptococcus pneumoniae dan Neisseria meningitidis diketahui
melepaskan polisakarida kapsuler selama pertumbuhan dalam jaringan. Bakteri ini
ditemukan dalam serum pasien dengan pneumonia pneumokokus dan dalam cairan
serebrospinal pasien dengan meningitis. Komponen-komponen sel bakteri yang
larut dalam dinding adalah antigen yang kuat dan melengkapi aktivator sehingga
mereka berkontribusi dengan cara utama untuk patologi yang diamati pada
penderita meningitis dan pneumonia.
Secara umum tahapan sistem kekebalan
tubuh terhadap mikroba adalah sebagai berikut:
Tahap pertama
Respons
inflamasi tubuh merupakan salah satu sel tubuh yang timbul sebagai akibat
invasi mikroba pada jaringan. Respons ini terdiri dari aktivitas sel-sel
inflamasi, antara lain sel leukosit (polimorfonuklear, limfosit, monosit), sel
makrofag, sel mast, sel natural killer, serta suatu sistem mediator
kimia yang kompleks baik yang dihasilkan oleh sel (sitokin) maupun yang
terdapat dalam plasma. Sel fagosit, mononuklear maupun polimorfonuklear
berfungsi pada proses awal untuk membunuh mikroba, dan mediator kimia dapat
meningkatkan fungsi ini. Mediator kimia akan berinteraksi satu dengan lainnya,
juga dengan sel radang seperti komponen sistem imun serta fagosit, baik
mononuklear maupun polimorfonuklear untuk memfagosit dan melisis mikroba.
Mediator tersebut antara lain adalah histamin, kinin/bradikinin, komplemen,
prostaglandin, leukotrien dan limfokin. Respons inflamasi ini bertujuan untuk
mengeliminasi dan menghambat penyebaran mikroba.
Histamin
yang dilepaskan sel mast akibat stimulasi anafilatoksin akan menyebabkan
vasodilatasi dan peningkatan permeabilitas vaskular untuk memfasilitasi
peningkatan aliran darah dan keluarnya sel radang intravaskular ke jaringan
tempat mikroba berada. Kinin/bradikinin adalah peptida yang diproduksi sebagai
hasil kerja enzim protease kalikrein pada kininogen. Mediator ini juga
menyebabkan vasodilatasi dan peningkatan permeabilitas pembuluh darah. Faktor
Hageman yang diaktifkan oleh karena adanya kerusakan pembuluh darah serta
endotoksin bakteri gram negatif, juga sel dalam menginduksi mediator kimia
lainnya.
Produk
aktivasi komplemen yang pada mulanya melalui jalur alternatif dapat
meningkatkan aliran darah, permeabilitas pembuluh darah, keinotaksis dan
fagositosis, serta hasil akhir aktivasi komplemen adalah lisis mikroba. Prostaglandin,
leukotrien dan fosfolipid lainnya yaitu mediator yang merupakan hasil metabolit
asam arakidonat dapat menstimulasi motilitas leukosit yang dibutuhkan untuk
memfagosit mikroba dan merangsang agregasi trombosit untuk memperbaiki
kerusakan pembuluh darah yang ada. Prostaglandin juga dapat bekerja sebagai
pirogen melalui pusat termoregulator di hipotalamus. Dikatakan bahwa panas juga
merupakan mekanisme sel tubuh, tetapi sukar dibuktikan. Mikroba tertentu memang
tidak dapat hidup pada suhu panas tetapi suhu tubuh yang tinggi akan memberikan
dampak yang buruk pada pejamu.
Protein
fase akut seperti C-reactive protein (CRP), protein yang mengikat
lipopolisakarida, protein amiloid A, transferin dan α1-antitripsin akan
dilepaskan oleh hati sebagai respons terhadap inflamasi. Peranannya dapat
sebagai stimulator atau inhibisi. Protein α1-antitripsin misalnya akan
menghambat protease yang merangsang produksi kinin. Transferin yang mempunyai
daya ikat terhadap besi, akan menghambat proliferasi dan pertumbuhan mikroba.
Protein yang mengikat lipopolisakarida akan menginaktifkan endotoksin bakteri
Gram negatif.
Limfokin,
yaitu sitokin yang dihasilkan limfosit, merupakan mediator yang kuat dalam
respons inflamasi. Limfokin ini dan sebagian diantaranya juga disekresi oleh
makrofag akan meningkatkan permeabilitas vaskular dan koagulasi, merangsang
produksi prostaglandin dan faktor kemotaksis, merangsang diferensiasi sel induk
hematopoietik dan meningkatkan pertumbuhan serta diferensiasi sel
hematopoietik, serta mengaktivasi neutrofil dan sel endotel. Sel radang yang
ada akan memfagosit mikroba, sedangkan monosit dan makrofag juga akan
memfagosit debris pejamu dan patogen yang tinggal sebagai hasil penyerangan
enzim neutrofil dan enzim lainnya. Fungsi makrofag akan ditingkatkan oleh
faktor aktivasi makrofag seperti komponen C3b, interferon γ dan faktor aktivasi
makrofag yang disekresi limfosit.
Tahapan kedua
Jika
mikroba berhasil melampaui mekanisme sel nonspesifik, terjadi tahapan kedua
berupa pertahanan spesifik yang dirangsang oleh antigen mikroba itu sendiri,
atau oleh antigen yang dipresentasikan makrofag. Tahapan ini terdiri atas
imunitas humoral dan imunitas selular.
Imunitas humoral yang diperankan oleh
antibodi yang dihasilkan oleh sel plasma sebagai hasil aktivasi antigen mikroba
terhadap limfosit B, akan menetralkan toksin yang dilepaskan mikroba sehingga
tidak menjadi toksik lagi. Antibodi juga akan menetralkan mikroba sehingga
tidak infeksius lagi. Antibodi juga bersifat sebagai opsonin, sehingga
memudahkan proses fagositosis mikroba. Antibodi juga berperan dalam proses
ADCC (Antibody Dependent Cell Cytotoxicity) baik oleh sel Tc maupun
sel NK sehingga terjadi lisis sel yang telah dihuni mikroba. Antibodi juga
dapat mengaktifkan komplemen untuk melisis mikroba. Imunitas selular yang
diperankan oleh limfosit T melalui limfokin yang dilepas sel T akan
meningkatkan produksi antibodi oleh sel plasma, fungsi sel fagosit untuk
memfagosit mikroba; dan sel NK untuk melisis sel yang dihuni virus (lihat Bab
3). Limfokin juga meningkatkan proliferasi dan diferensiasi sel prekursor Tc
serta fungsi sel Tc untuk melisis sel yang dihuni mikroba. Inteleukin (IL)- 2,
IL-12 dan IFN-γ meningkatkan imunitas selular. Imunitas selular adalah
mekanisme utama tubuh untuk terminasi infeksi mikroba intraselular seperti
infeksi virus, parasit dan bakteri intraselular.
Tahapan Akhir
Tahapan
terakhir ini terdiri atas peningkatan respons imun baik melalui aktivasi
komplemen jalur klasik maupun peningkatan kemotaksis, opsonisasi dan fagositosis.
Sel makrofag dan limfosit T terus memproduksi faktor yang selanjutnya akan
meningkatkan lagi respons inflamasi melalui ekspresi molekul adesi pada endotel
serta merangsang kemotaksis, pemrosesan antigen, pemusnahan intraselular,
fagositosis dan lisis, sehingga infeksi dapat teratasi.
Respons
imun yang terkoordinasi yang melibatkan sel T, antibodi, sel makrofag, sel PMN,
komplemen dan pertahanan nonspesifik lainnya akan terjadi pada kebanyakan
penyakit infeksi.
Mekanisme Respon Tubuh terhadap Serangan Virus
Virus
berbeda dengan agen penyebab infeksi lainnya dalam hal struktur dan biologi,
khususnya reproduksi. Walaupun virus membawa informasi genetik didalam DNA atau
RNA, tetapi ada kekurangan sistem sintesis yang diperlukan untuk memproses
informasi ini kedalam materi virus baru. Replikasi baru terjadi setelah virus
menginfeksi sel inang yang kemudian mengendalikan sel inang untuk melakukan
transkripsi dan/atau translasi informasi genetik demi kelangsungan hidup virus.
Virus dapat menginfeksi setiap bentuk kehidupan sehingga sering menyebabkan
penyakit yang diantaranya berakibat cukup serius. Beberapa virus dapat
memasukkan informasi genetiknya kedalam genom manusia kemudian menyebabkan
kanker. Permukaan luar partikel virus adalah bagian yang pertamakali
mengadakan kontak dengan membran dari sel inang. Hal yang penting untuk
diketahui untuk dapat mengerti bagaimana proses virus dapat menginfeksi sel
inang adalah dengan mempelajari struktur dan fungsi dari permukaan luar
partikel virus. Secara umum, virus yang tidak beramplop (virus yang telanjang)
resisten hidup dialam bebas, bahkan mereka tahan terhadap asam empedu saat
menginfeksi saluran cerna. Virus yang beramplop lebih rentan terhadap
dipengaruhi oleh lingkungan seperti kekeringan, asiditas cairan lambung dan
empedu. Perbedaan dalam hal kerentanan ini yang mempengaruhi cara
penularan virus.
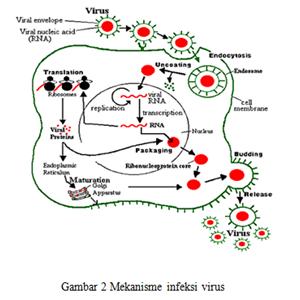
Infeksi
virus terhadap sel inang melewati beberapa tahap, yaitu virus menyerang sel
inang, lalu melakukan penetrasi yang merupakan proses pemasukan materi genetik virus
kedalam sel inang dan selanjutnya tahap uncoating yang ditunjukan
pada gambar 1.
Siklus
hidup yang dialami virus saat menginfeksi sel inang, yaitu sekali virus berada
didalam sitoplasma sel inang maka dia tidak infeksius lagi. Setelah terjadi
fusi antara virus dan membramn sel inang, atau difagosit dalam bentuk fagosom,
maka partikel virus dibawa ke sitoplasma melalui plasma membran. Pada tahap ini
amplop dan/atau kapsid akan terkuak nukleus virus akan terurai. Sekarang virus
tidak infeksius lagi dan ini disebut eclipse phase. Keadaan ini menetap
sampai terbentuk partikel virus baru melalui replikasi. Asam nukleat sendiri
yang menentukan bagaimana cara replikasi berlangsung. Pertama-tama virus harus
membentuk messenger RNA (mRNA). Virus hanya mempunyai salah
satu asam nukleat yaitu RNA atau DNA dan tidak pernah kedua-duanya. Asam
nukleat tampil sebagai single atau double strandad dalam
bentuk linier (DNA dan RNA) atau sirkuler (DNA). Genom dari virus terdapat
dalam satu atau beberapa molekul dari asam nukleat.
Dengan
diversitas ini maka tidak heran bila proses replikasi dari tiap virus berbeda.
Pada virus DNA, mRNA dapat dibentuk sendiri oleh virus dengan cara menggunakan
RNA polimerase dari sel inang, kemudian langsung mentranskrip kode genetik yang
berada pada DNA virus. Sedangkan virus RNA tidak dapat dengan cara ini, karena
tidak ada polymerase dari sel inang yang sesuai. Oleh karena itu
untuk melakukan transkripsi maka virus harus menyediakan sendiri
polimerasenya yang dapat diperoleh dari nukleokapsid atau disintesa setelah
infeksi.
Virus RNA
memproduksi mRNA dengan beberapa cara yang berbeda. Pada virus dsRNA,
satu strand yang pertama ditranskrip oleh polimerase virus menjadi
mRNA. Pada ssRNA terdapat tiga rute yang jelas berbeda dalam pembentukan mRNA
yaitu:
·
Bila single strand mempunyai
konfigurasi positive sense (misalnya mempunyai sekuen basa yang sama
seperti yang dibutuhkan pada saat translasi), maka konfigurasi ini dapat
langsung dipergunakan sebagai mRNA.
·
Bila mempunyai konfigurasi negative
sense, maka pertama-tama harus diterjemahkan (transcribe) dengan memgunakan
polimerase dari virus kedalam positive sense strand yang
kemudian bertindak sebagai mRNA.
Retrovirus
mempunyai pola yang berbeda. Pertama-tama positive sense ssRNA
oleh reverse transcriptase (enzim dari virus, terdapat dalam
nukleokapsid) diubah menjadi negative sense ssDNA. Setelah
terbentuk dsDNA kemudian akan memasuki nukleus dan kemudian berintegrasi
dengan genom sel inang dan selanjutnya sel inang membentuk mRNA virus.
Tahapan
selanjutnya yaitu, mRNA virus kemudian ditranslasi kedalam sitoplasma sel inang
untuk menghasilkan protein yang dibutuhkan virus. Sekali mRNA virus terbentuk
maka akan ditanslasi dengan memanfaatkan ribosom dari sel inang untuk
mensintesa protein yang dibutuhkan virus dan ditunjukkan pada Gambar 3. RNA
virus biasanya monocistronic(mempunyai single coding region) dapat
mengubah mRNA dari ribosom sel inang untuk menghasilkan protein yang lebih
‘disukai’. Pada fase awal diproduksi protein yang diperlukan untuk replikasi
asam nukleat virus seperti enzim dan molekul regulator. Pada fase
selanjutnya diproduksi protein yang penting unutk pembentukan kapsid. Virus
dengan genom single nucleic acid molecule mentranslasi poli protein
yang multifungsi, kemudian akan dipecah secara enzimatik. Sedangkan virus yang
genomnya tersebar didalam beberapa molekul, maka akan terbentuk beberapa
macam mRNA yang masing-masing akan membuat protein. Setelah translasi protein
dapat diglikosilasi kembali dengan menggunakan enzim sel inang.
Virus juga
harus mereplikasi asam nukleatnya untuk pembentukan kapsid baru berarti
memerlukan produksi molekul tambahan. Oleh karena itu virus harus mereplikasi
asam nukleat sehingga dapat menyediakan materi genetik yang kemudian akan
dibungkus oleh kapsid tersebut. Pada virus positive sense ssRNA
seperti poliovirus, polimerase yang ditranslasi dari template mRNA virus
menghasilkan negative sense RNA yang selanjutnya ditranskripsi lebih
banyak positif ssRNA. Siklus transkripsi ini terus berlangsung menghasilkan
strand positif dalam jumlah yang besar, yang kemudian dikemas dengan
menggunakan protein yang telah dibentuk sebelumnya dari mRNA untuk membentuk
partikel virus yang baru. Untuk virus negative sense ssRNA
(misalnya virus rabies) transkripsi oleh polimerase virus akan
menghasilkan positive sense ssRNA yang kemudian akan
meghasilkan negative sense mRNA yang baru.
Replikasi
ini terjadi dalam sitoplasma sel inang, sedangkan pada virus lainnya
seperti campak dan influensa replikasi terjadi di inti sel sehingga
sejumlah besar negative sense RNA akan ditranskripsi membentuk
partikel baru. Replikasi pada inti sel inang juga terjadi pada virus
dsRNA seperti rotavirus yang kemudian akan memproduksi positive
sense RNA seperti diatas. Yang kemudian akan bertindak sebagai template
pada partikel subviral untuk memsintesa negative senseRNA yang baru guna
memperbaiki kondisi double stranded. Replikasi virus DNA terjadi di
inti sel inang kecuali poxvirus yang terjadi di sitoplasma Virus DNA membentuk
kompleks dengan histon dari sel inang untuk menghasilkan struktur yang stabil.
 Pada virus
herpes, mRNA ditranslasi dalam sitoplasma
menghasilkan polymerase DNA yang penting untuk sintesa DNA yang baru.
Adenovirus menggunakan baik enzim dari sel inang maupun virus untuk
kepentingan ini. Sedangkan retrovirus mensintesa RNA virus baru di inti sel
inang. Polimerase RNA sel inang ditranskrip dari DNA virus yang sudah
berintegrasi dengan genom sel inang. Virus hepatitis B (suatu virus dsDNA)
secara unik menggunakan ssRNA (sebagai perantara) yang kemudian ditranskrip
untuk menghasilkan DNA baru. Retrovirus dan virus hepatitis B merupakan
virus-virus yang mempunyai aktifitas reverse transkriptase.
Pada virus
herpes, mRNA ditranslasi dalam sitoplasma
menghasilkan polymerase DNA yang penting untuk sintesa DNA yang baru.
Adenovirus menggunakan baik enzim dari sel inang maupun virus untuk
kepentingan ini. Sedangkan retrovirus mensintesa RNA virus baru di inti sel
inang. Polimerase RNA sel inang ditranskrip dari DNA virus yang sudah
berintegrasi dengan genom sel inang. Virus hepatitis B (suatu virus dsDNA)
secara unik menggunakan ssRNA (sebagai perantara) yang kemudian ditranskrip
untuk menghasilkan DNA baru. Retrovirus dan virus hepatitis B merupakan
virus-virus yang mempunyai aktifitas reverse transkriptase.
Stadium
akhir dari replikasi adalah penyusunan dan pelepasan parikel virus baru.
Penyusunan virus baru melibatkan gabungan dari asam nukleat yang telah
direplikasi dengan kapsomer yang baru disintesa untuk kemudian membentuk
nukleokapsid baru. Aktifitas ini terjadi di sitoplasma atau di inti sel inang.
Amplop dari virus melalui beberapa tahapan sebelum dilepaskan. Protein amplop
dan glikoprotein yang ditranslasi dari mRNA virus didisipkan pada membran sel
inang (biasanya membrana plasma). Nukleokapsid yang muda ini bergabung dengan
membran secara spesifik melalui glikoprotein dan menbentuk tonjolan. Virus baru
memerlukan membran dari sel inang ditambah dengan molekul dari virus untuk
membentuk amplop. Enzim dari virus seperti muraminidase pada virus influensa
ikut berperan dalam proses ini. Enzim dari sel inang (seperti protease seluler)
dapat memecah protein amplop yang besar, suatu proses yang diperlukan
dimana virus muda sangat infeksius. Pada virus herpes terjadi proses yang sama.
Pelepasan virus yang sudah beramplop tidak harus disertai dengan kematian sel,
jadi sel inang yang sudah terinfeksi dapat terus menghasilkan protein virus
dalam waktu yang lama. Insersi molekul virus kedalam membran sel inang membuat
sel inang berbeda secara antigenik. Respon imun ekspresi antigen ini yang
menjadi dasar perkembangan terapi anti virus.
Pada
respon innate terhadap patogen intraseluler, seperti virus, sasaran
utama adalah sel-sel yang sudah terinfeksi. Sel terinfeksi virus tertentu
dikenali oleh limfosit non-spesifik, disebut sel natural killer (NK).
Sesuai dengan namanya, sel NK mengakibatkan kematian sel yang terinfeksi
dengan menginduksi sel terinfeksi menuju apoptosis. Sel NK juga membunuh
sel kanker tertentu (in vitro) dan melengkapi dengan mekanisme menghancurkan
sel sebelum sel berkembang menjadi tumor. Sel normal (tidak terinfeksi dan
tidak ganas) mengandung molekul permukaan yang melindungi terhadap serangan sel
NK. Respon antivirus lain dimulai dalam sel yang terinfeksi sendiri. Sel
terinfeksi virus ini memproduksi interferon-α (IFN-α) yang disekresi ke dalam
ruang ekstraseluler, dimana akan terikat pada permukaan sel yang tidak
terinfeksi sehingga kebal terhadap infeksi berikutnya. Cara kerja
interferon ini adalah dengan cara mengaktivasi suatu sinyal transduction
pathway dengan akibat phosphorilasi yang diikuti translasi faktor elF2.
Sel yang mengalami respons ini tidak dapat mensintesa protein virus yang
diperlukan untuk replikasi virus.
Respon
imun terhadap serangan virus melibatkan interferon. Interferon merupakan
sitokin yang mengatur aktivitas semua komponen sistem imun,
merupakan bagian dari sistem imun
non-spesifik yang timbul pada tahap awal
infeksi virus sebelum timbulnya reaksi dari sistem imun
spesifik. Interferon gamma (IFN-γ) dihasilkan oleh
sel T yang telah teraktivasi dan sel
NK, sebagai reaksi terhadap antigen (termasuk
antigen virus dalam derajat rendah) atau
sebagai akibat stimulasi limfosit oleh
mitogen. IFN-γ meningkatkan ekspresi molekul
MHC-II pada Antigen Presenting Cell (APC)
yang kemudian akan meningkatkan presentasi
antigen pada sel T helper. IFN-γ juga
dapat mengaktifkan kemampuan makrofag untuk melawan
infeksi virus (aktivitas virus intrinsik) dan membunuh
sel lain yang telah terinfeksi (aktivitas virus ekstrinsik)
(Ianaro 2000).
Mekanisme Respon Tubuh terhadap Serangan Bakteri
Bakteri
adalah kelompok organisme yang tidak memiliki membran inti. Beberapa kelompok
bakteri dikenal sebagai agen penyebab infeksi dan penyakit. Bakteri dapat
ditemukan di hampir semua tempat seperti di tanah, air, udara, dalam simbiosis
dengan organisme lain maupun sebagai agen parasit (patogen), bahkan dalam tubuh
manusia. Respon imun terhadap sebagian besar antigen seperti bakteri ini hanya
dimulai bila antigen telah ditangkap dan diproses serta dipresentasikan oleh
sel APC (Antigen Presenting Cell).
Keberhasilan
bakteri masuk ke dalam sitoplasma sel bergantung pada kemampuannya untuk
menghindar dari respon imun. Infeksi bakteri akan berbeda sesuai dengan sistem
kerja dari bakteri tersebut. Dimana dalam hal ini dipaparkan infeksi bakteri
ekstraseluler dan interaseluler beserta mekanisme pertahanan tubuh manusia
(Munasir 2001).
Infeksi
bakteri berbeda dengan infeksi virus. Respons imun terhadap bakteri ada dua
yaitu, ekstraselular dan intraselular.
1.
Respons imun terhadap bakteri ekstraselular
Bakteri
ekstraselular dapat menimbulkan penyakit melalui beberapa mekanisme yaitu:
·
Merangsang reaksi inflamasi yang menyebabkan
destruksi jaringan di tempat infeksi. Sebagai contoh misalnya kokus piogenik
yang sering menimbulkan infeksi supuratif yang hebat.
 ·
Produksi toksin yang menghasilkan berbagai
efek patologik. Toksin dapat berupa endotoksin dan eksotoksin. Endotoksin yang
merupakan komponen dinding bakteri adalah suatu lipopolisakarida yang merupakan
stimulator produksi sitokin yang kuat, suatu ajuvan serta aktifator poliklonal
sel limfosit B. Sebagian besar eksotoksin mempunyai efek sitotoksik dengan
mekanisme yang belum jelas benar. Sebagai contoh toksin difteri menghambat
sintesis protein secara enzimatik serta menghambat faktor elongasi-2 yang
diperlukan untuk sintesis semua peptida. Toksin kolera merangsang sintesis AMP
siklik (cAMP) oleh sel epitel usus yang menyebabkan sekresi aktif klorida,
kehilangan cairan serta diare yang hebat. Toksin tetanus merupakan suatu
neurotoksin yang terikat
motor endplate pada neuromuscular junction yang
menyebabkan kontraksi otot persisten yang sangat fatal bila mengenai otot
pernapasan. Toksin Clostridium dapat menyebabkan nekrosis jaringan
yang dapat menghasilkan gas gangren. Respon imun terhadap bakteri ekstraselular
ditujukan untuk eliminasi bakteri serta netralisasi efek toksin
·
Produksi toksin yang menghasilkan berbagai
efek patologik. Toksin dapat berupa endotoksin dan eksotoksin. Endotoksin yang
merupakan komponen dinding bakteri adalah suatu lipopolisakarida yang merupakan
stimulator produksi sitokin yang kuat, suatu ajuvan serta aktifator poliklonal
sel limfosit B. Sebagian besar eksotoksin mempunyai efek sitotoksik dengan
mekanisme yang belum jelas benar. Sebagai contoh toksin difteri menghambat
sintesis protein secara enzimatik serta menghambat faktor elongasi-2 yang
diperlukan untuk sintesis semua peptida. Toksin kolera merangsang sintesis AMP
siklik (cAMP) oleh sel epitel usus yang menyebabkan sekresi aktif klorida,
kehilangan cairan serta diare yang hebat. Toksin tetanus merupakan suatu
neurotoksin yang terikat
motor endplate pada neuromuscular junction yang
menyebabkan kontraksi otot persisten yang sangat fatal bila mengenai otot
pernapasan. Toksin Clostridium dapat menyebabkan nekrosis jaringan
yang dapat menghasilkan gas gangren. Respon imun terhadap bakteri ekstraselular
ditujukan untuk eliminasi bakteri serta netralisasi efek toksin
Imunitas Alamiah terhadap Bakteri Ekstraselular
Respon
imun alamiah terhadap bakteri ekstraselular terutama melalui mekanisme fagositosis
oleh neutrofil, monosit serta makrofag jaringan. Resistensi bakteri terhadap
fagositosis dan penghancuran dalam makrofag menunjukkan virulensi bakteri.
Aktivasi komplemen tanpa adanya antibodi juga memegang peranan penting dalam
eliminasi bakteri ekstraselular. Lipopolisakarida (LPS) dalam dinding bakteri
gram negatif dapat mengaktivasi komplemen jalur alternatif tanpa adanya
antibodi. Salah satu hasil aktivasi komplemen ini yaitu C3b mempunyai efek
opsonisasi bakteri serta meningkatkan fagositosis. Selain itu terjadi lisis
bakteri melaluimembrane attack complex (MAC) serta beberapa hasil
sampingan aktivasi komplemen dapat menimbulkan respon inflamasi melalui
pengumpulan serta aktivasi leukosit. Endotoksin yang merupakan LPS merangsang
produksi sitokin oleh makrofag serta sel lain seperti endotel vaskular.
Beberapa jenis sitokin tersebut antara lain tumour necrosis
factor (TNF), IL-1, IL-6 serta beberapa sitokin inflamasi dengan berat
molekul rendah yang termasuk golongan IL-8. Fungsi fisiologis yang utama dari
sitokin yang dihasilkan oleh makrofag adalah merangsang inflamasi non-spesifik
serta meningkatkan aktivasi limfosit spesifik oleh antigen bakteri. Sitokin
akan menginduksi adhesi neutrofil dan monosit pada endotel vaskular pada tempat
infeksi yang diikuti migrasi, akumulasi lokal serta aktivasi sel inflamasi.
Kerusakan
jaringan yang terjadi adalah akibat efek samping mekanisme pertahanan untuk
eliminasi bakteri tersebut. Sitokin juga merangsang demam dan sintesis protein
fase akut. Banyak fungsi sitokin yang sama yaitu sebagai co-stimulator sel
limfosit T dan B yang menghasilkan mekanisme amplifikasi untuk imunitas
spesifik. Sitokin dalam jumlah besar atau produknya yang tidak terkontrol dapat
membahayakan tubuh serta berperan dalam menifestasi klinik infeksi bakteri
ekstraselular. Yang paling berat adalah gejala klinis oleh infeksi bakteri gram
negatif yang menyebabkan disseminated intravascular
coagulation (DIC) yang progresif serta shock septik
atau shock endotoksin. Sitokin TNF adalah mediator yang paling
berperan pada shock endotoksin ini.
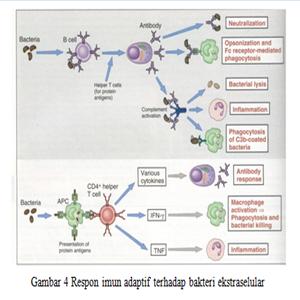
Imunitas Spesifik terhadap Bakteri Ekstraselular
Kekebalan
humoral mempunyai peran penting dalam respon kekebalan spesifik terhadap
bakteri ekstraselular. Lipopolisakarida merupakan komponen yang paling
imunogenik dari dinding sel atau kapsul mikroorganisme serta merupakan antigen
yang thymus independent. Antigen ini dapat langsung merangsang sel
limfosit B yang menghasilkan imunoglobin (Ig)M spesifik yang kuat. Selain itu
produksi IgG juga dirangsang yang mungkin melalui mekanisme
perangsangan isotype switching rantai berat oleh sitokin. Respon sel
limfosit T yang utama terhadap bakteri ekstraselular melalui sel TCD4 yang
berhubungan dengan molekul MHC kelas II yang mekanismenya telah dijelaskan sebelumnya.
Sel TCD4 berfungsi sebagai sel penolong untuk merangsang pembentukan antibodi,
aktivasi fungsi fagosit dan mikrobisid makrofag. Ada 3 mekanisme efektor yang
dirangsang oleh IgG dan IgM serta antigen permukaan bakteri, yaitu:
·
Opsonisasi bakteri oleh IgG serta peningkatan
fagositosis dengan mengikat reseptor Fc pada monosit, makrofag dan neutrofil.
Antibodi IgG dan IgM mengaktivasi komplemen jalur klasik yang menghasilkan C3b
dan iC3b yang mengikat reseptor komplemen spesifik tipe 1 dan tipe 3 dan
selanjutnya terjadi peningkatan fagositosis. Pasien defisiensi C3 sangat rentan
terhadap infeksi piogenik yang hebat.
·
Netralisasi toksin bakteri oleh IgM dan IgG
untuk mencegah penempelan terhadap sel target serta meningkatkan fagositosis
untuk eliminasi toksin tersebut.
Aktivasi
komplemen oleh IgM dan IgG untuk menghasilkan mikrobisid MAC serta pelepasan
mediator inflamasi akut.
1.
Respons Imun terhadap Bakteri Intraselular
Sejumlah
bakteri dan semua virus serta jamur dapat lolos dan mengadakan replikasi di
dalam sel pejamu. Yang paling patogen di antaranya adalah yang
resisten terhadap degradasi dalam makrofag.
Imunitas
Alamiah terhadap Bakteri Intraselular
Mekanisme
terpenting imunitas alamiah terhadap mikroorganisme intraselular adalah
fagositosis. Akan tetapi bakteri patogen intraselular relatif resisten terhadap
degradasi dalam sel fagosit mononuklear. Oleh karena itu mekanisme kekebalan
alamiah ini tidak efektif dalam mencegah penyebaran infeksi sehingga sering
menjadi kronik dan eksaserbasi yang sulit diberantas.
Imunitas
Spesifik terhadap Bakteri Intraselular
Respon
imun spesifik terhadap bakteri intraselular terutama diperankan
oleh cell mediated immunity (CMI). Mekanisme imunitas ini
diperankan oleh sel limfosit T tetapi fungsi efektornya untuk eliminasi bakteri
diperani oleh makrofag yang diaktivasi oleh sitokin yang diproduksi oleh sel T
terutama interferon-α (IFN-α). Respon imun ini analog dengan reaksi
hipersensitivitas tipe lambat. Antigen protein intraselular merupakan stimulus
kuat sel limfosit T. Beberapa dinding sel bakteri mengaktivasi makrofag secara
langsung sehingga mempunyai fungsi sebagai ajuvan. Misalnya muramil dipeptida
pada dinding sel mikrobakteria. Telah disebutkan sebelumnya bahwa fungsi sel
limfosit T pada CMI adalah produksi sitokin terutama IFN-α. Sitokin IFN-α ini
akan mengaktivasi makrofag termasuk makrofag yang terinfeksi untuk membunuh
bakteri. Beberapa bakteri ada yang resisten sehingga menimbulkan stimulasi
antigen yang kronik. Keadaan ini akan menimbulkan pengumpulan lokal makrofag
yang teraktivasi yang membentuk granuloma sekeliling mikroorganisme untuk
mencegah penyebarannya.
Reaksi
inflamasi seperti ini berhubungan dengan nekrosis jaringan serta fibrosis yang
luas yang menyebabkan gangguan fungsi yang berat. Jadi kerusakan jaringan ini
disebabkan terutama oleh respon imun terhadap infeksi oleh beberapa bakteri
intraselular. Contoh yang jelas dalam hal ini adalah infeksi mikobakterium.
Mikobakterium tidak memproduksi toksin atau enzim yang secara langsung merusak
jaringan yang terinfeksi. Paparan pertama terhadap Mycobacterium
tuberculosis akan merangsang inflamasi selular lokal dan bakteri
mengadakan proliferasi dalam sel fagosit. Sebagian ada yang mati dan sebagian
ada yang tinggal dormant. Pada saat yang sama, pada individu yang terinfeksi
terbentuk imunitas sel T yang spesifik. Setelah terbentuk imunitas, reaksi
granulomatosa dapat terjadi pada lokasi bakteri persisten atau pada paparan
bakteri berikutnya. Jadi imunitas perlindungan dan reaksi hipersensitif yang
menyebabkan kerusakan jaringan adalah manifestasi dalam respon imun spesifik
yang sama.
Netralisasi toksin
Infeksi
bakteri Gram negatif dapat menyebabkan pengeluaran endotoksin yang akan
menstimulasi makrofag. Stimulasi yang berlebihan terhadap makrofag akan
menghasilkan sejumlah sitokin seperti IL-1, IL-6 dan TNF. Proses ini akan
memacu terjadinya reaksi peradangan yang menyebabkan kerusakan sel, hipotensi,
aktivasi sistem koagulasi, gagal organ multipel dan berakhir dengan kematian.
Antibodi yang mengandung reseptor sitokin dan antagonisnya, berperan dalam
menghilangkan sejumlah sitokin dalam sirkulasi dan mencegah sitokin berikatan
pada sel target.
Antibodi
yang beredar dalam sirkulasi akan menetralisasi molekul antifagositik dan
eksotoksin lainnya yang diproduksi bakteri. Mekanisme netralisasi antibodi
terhadap bakteri terjadi melalui dua cara. Pertama, melalui kombinasi antibodi
di dekat lokasi biologi aktif infeksi yaitu secara langsung menghambat reaksi
toksin dengan sel target. Kedua, melalui kombinasi antibodi yang terletak jauh
dari lokasi biologi aktif infeksi yaitu dengan mengubah konformasi alosterik
toksin agar tidak dapat bereaksi dengan sel target. Dengan ikatan kompleks
bersama antibodi, toksin tidak dapat berdifusi sehingga rawan terhadap
fagositosis, terutama bila ukuran kompleks membesar karena deposisi komplemen
pada permukaan bakteri akan semakin bertambah.
Opsonisasi
Opsonisasi
adalah pelapisan antigen oleh antibodi, komplemen, fibronektin, yang berfungsi
untuk memudahkan fagositosis. Opsonisasi ada dua yaitu opsonisasi yang tidak
tergantung antibodi dan yang ditingkatkan oleh antibodi. Pada opsonisasi yang
tidak tergantung antibodi, protein pengikat manose dapat terikat pada manose
terminal pada permukaan bakteri, dan akan mengaktifkan C1r dan C1s serta
berikatan dengan C1q. Proses tersebut akan mengaktivasi komplemen pada jalur
klasik yang dapat berperan sebagai opsonin dan memperantarai fagositosis.
Lipopolisakarida (LPS) merupakan endotoksin yang penting pada bakteri Gram
negatif. Sel ini dapat dikenal oleh tiga kelas molekul reseptor. Sedangkan
opsonisasi yang ditingkatkan oleh antibodi adalah bakteri yang resisten
terhadap proses fagositosis akan tertarik pada sel PMN dan makrofag bila telah
diopsonisasi oleh antibodi. Dalam opsonisasi terdapat sinergisme antara
antibodi dan komplemen yang diperantarai oleh reseptor yang mempunyai afinitas
kuat untuk IgG dan C3b pada permukaan fagosit, sehingga meningkatkan pengikatan
di fagosit. Efek augmentasi dari komplemen berasal dari molekul IgG yang dapat
mengikat banyak molekul C3b, sehingga meningkatkan jumlah hubungan ke
makrofag (bonus effect of multivalency). Meskipun IgM tidak terikat secara
spesifik pada makrofag, namun merangsang adesi melalui pengikatan komplemen.
Antibodi
akan menginisiasi aksi berantai komplemen sehingga lisozim serum dapat masuk ke
dalam lapisan peptidoglikan bakteri dan menyebabkan kematian sel. Aktivasi
komplemen melalui penggabungan dengan antibodi dan bakteri juga menghasilkan
anfilaktoksin C3a dan C5a yang berujung pada transudasi luas dari komponen
serum, termasuk antibodi yang lebih banyak, dan juga faktor kemotaktik
terhadap neutrofil untuk membantu fagositosis.
Sel PMN merupakan fagosit yang
predominan dalam sirkulasi dan selalu tiba di lokasi infeksi lebih cepat dari
sel lain, karena sel PMN tertarik oleh sinyal kemotaktik yang dikeluarkan oleh
bakteri, sel PMN lain, komplemen atau makrofag lain, yang lebih dahulu tiba di
tempat infeksi. Sel PMN sangat peka terhadap semua faktor kemotaktik. Sel PMN
yang telah mengalami kemotaktik selanjutnya akan melakukan adesi pada dinding
sel bakteri, endotel maupun jaringan yang terinfeksi. Kemampuan adesi PMN pada
permukaan sel bakteri akan bertambah kuat karena sinyal yang terbentuk pada
proses adesi ini akan merangsang ekspresi Fc dan komplemen pada permukaan sel.
Sel PMN juga akan melakukan proses diapedesis agar dapat menjangkau bakteri
yang telah menginfeksi.
Proses penelanan bakteri oleh fagosit
diawali dengan pembentukan tonjolan pseudopodia yang berbentuk kantong fagosom
untuk mengelilingi bakteri, sehingga bakteri akan terperangkap di dalamnya,
selanjutnya partikel granular di dalam fagosom akan mengeluarkan berbagai enzim
dan protein untuk merusak dan menghancurkan bakteri tersebut.
Mekanisme pemusnahan bakteri oleh
enzim ini dapat melalui proses oksidasi maupun nonoksidasi, tergantung pada
jenis bakteri dan status metabolik pada saat itu. Oksidasi dapat berlangsung
dengan atau tanpa mieloperoksidase. Proses oksidasi dengan mieloperoksidase
terjadi melalui ikatan H2O2 dengan Fe yang terdapat pada mieloperoksidase.
Proses ini menghasilkan komplek enzim-subtrat dengan daya oksidasi tinggi dan
sangat toksik terhadap bakteri, yaitu asam hipoklorat (HOCl).
Proses
oksidasi tanpa mieloperoksidase berdasarkan ikatan H2O2 dengan superoksida
dan radikal hidroksil namun daya oksidasinya rendah. Proses nonoksidasi
berlangsung dengan perantaraan berbagai protein dalam fagosom yaitu
flavoprotein, sitokrom-b, laktoferin, lisozim, kaptensin G dan
difensin. Pada proses pemusnahan bakteri, pH dalam sel fagosit dapat
menjadi alkalis. Hal ini terjadi karena protein yang bermuatan positif dalam pH
yang alkalis bersifat sangat toksik dan dapat merusak lapisan lemak dinding
bakteri Gram negatif. Selain itu, bakteri juga dapat terbunuh pada saat pH dalam
fagosom menjadi asam karena aktivitas lisozim. Melalui proses ini PMN
memproduksi antibakteri yang dapat berperan sebagai antibiotika alami (natural
antibiotics).
Sistem imun sekretori
Permukaan
mukosa usus mempunyai mekanisme pertahanan spesifik antigen dan nonspesifik.
Mekanisme nonspesifik terdiri dari peptida antimikrobial yang diproduksi oleh
neutrofil, makrofag dan epitel mukosa. Peptida ini akan menyebabkan lisis
bakteri melalui disrupsi pada permukaan membran. Imunitas spesifik diperantarai
oleh IgA sekretori dan IgM, dengan dominasi IgA1 pada usus bagian awal dan IgA2
pada usus besar. Antibodi IgA mempunyai fungsi proteksi dengan cara
melapisi (coating) virus dan bakteri dan mencegah adesi pada sel
epitel di membran mukosa. Reseptor Fc dari kelas Ig mempunyai afinitas tinggi
terhadap neutrofil dan makrofag dalam proses fagositosis. Apabila agen infeksi
berhasil melewati barier IgA, maka lini pertahanan berikutnya adalah IgE.
Adanya
kontak antigen dengan IgE akan menyebabkan pelepasan mediator yang menarik agen
respons imun dan menghasilkan reaksi inflamasi akut. Adanya peningkatan
permeabilitas vaskular yang disebabkan oleh histamin akan menyebabkan
transudasi IgG dan komplemen, sedangkan faktor kemotaktik terhadap neutrofil
dan eosinofil akan menarik sel efektor yang diperlukan untuk mengatasi
organisme penyebab infeksi yang telah dilapisi oleh IgG spesifik dan C3b.
Penyatuan kompleks antibodi-komplemen pada makrofag akan menghasilkan faktor
yang memperkuat permeabilitas vaskular dan proses kemotaktik. Apabila organisme
yang diopsonisasi terlalu besar untuk difagosit, maka fagosit dapat mengatasi
organisme tersebut melalui mekanisme ekstraseluler,
yaitu Antibody-Dependent Cellular Cytotoxicity (ADCC).
Terminologi Sitokin
Sitokin merupakan protein-protein kecil yang berfungsi sebagai mediator dan
pengatur immunitas, inflamasi dan hematopoesis. Sitokin disekresikan oleh
sel-sel tertentu dari sistem kekebalan tubuh yang membawa sinyal antara sel-sel
lokal sehingga memiliki efek pada sel lain. Sitokin dihasilkan sebagai respon
terhadap stimulus sistem imun. Sitokin bekerja dengan mengikat
reseptor-reseptor membran spesifik, yang kemudian membawa sinyal ke sel melalui
tirosine kinase (second messanger). Sitokina berfungsi sebagai sinyal interseluler
yang mengatur hampir semua proses biologi penting seperti halnya aktivasi,
pertumbuhan, proliferasi, diferensiasi, proses inflamasi sel, imunitas, serta
pertahanan jaringan ataupun morfogenesis. Sitokina mempunyai berat molekul
rendah sekitar 8-40 kilo dalton, di samping kadarnya juga sangat rendah.
Klasifikasi sel Sitokin
Sitokin
adalah nama umum dari hasil sekresi sel tertentu, nama yang lain diantaranya
limfokin (dihasilkan limfosit), monokin (sitokin yang dihasilkan monosit),
kemokin (sitokin dengan aktivitas kemotaktik), dan interkulin (sitokin yang
dihasilkan oleh satu leukosit dan bereaksi pada leukosit lain). Sitokina
biasanya diproduksi oleh sel sebagai respon terhadap rangsangan. Sitokina yang
dibentuk segera dilepas dan tidak disimpan di dalam sel. Satu sitokina dapat
bekerja terhadap beberapa jenis sel dan dapat menimbulkan efek melalui berbagai
mekanisme. Setiap jenis sitokin dihasilkan oleh sel berbeda dan digunakan pada
sel target yang berbeda juga sehingga fungsinya pun akan berbeda.
 Anda sedang membaca artikel berjudul
Anda sedang membaca artikel berjudul 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
0 komentar:
Posting Komentar